ハリー・ゴビエ・シーリーは恐竜を竜盤類(「トカゲ寄棟」恐竜)と鳥盤類(「鳥寄棟」恐竜)の2つのグループに分けました。どちらの命令にも、おそらく中期三畳紀のある時期に住んでいた共通の祖先がいました。鳥は竜盤類の恐竜クレードに属しています。
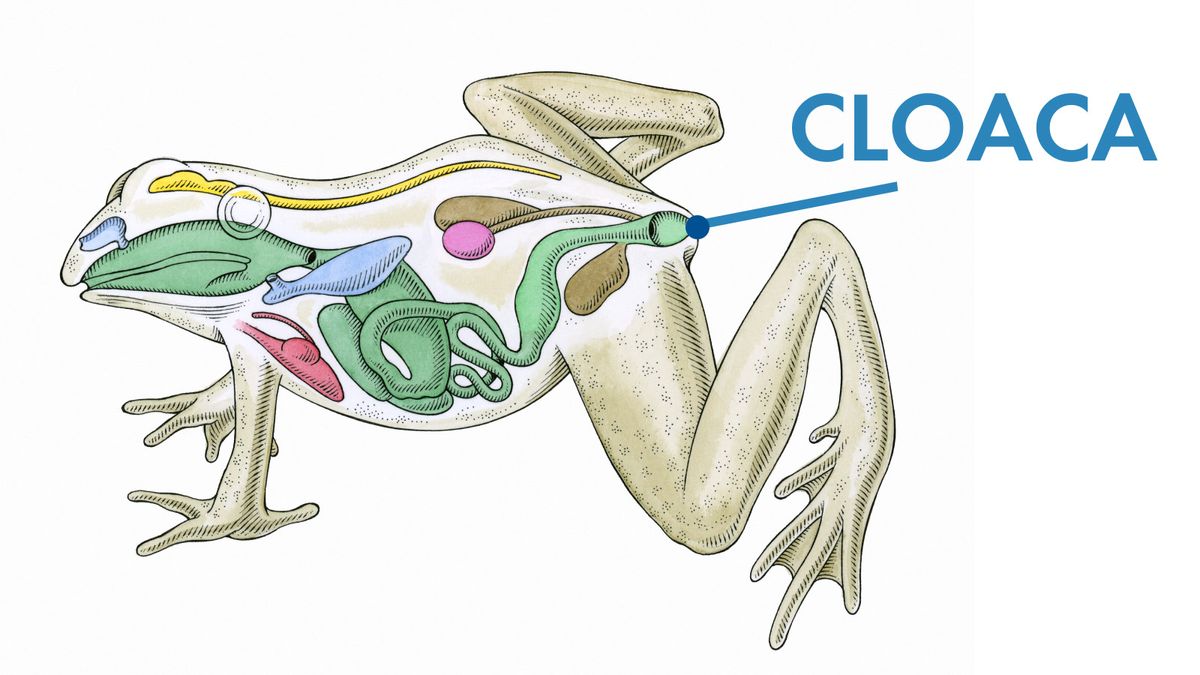
すべての陸上動物と同様に、骨盤の両側に3つの骨がありました。左右の腸骨(単数:腸骨)が仙骨の脊椎をしっかりとつかみました。左右の恥骨(単数:恥骨)は腸骨の下に伸びていました。左右のイスキア(単数:坐骨)は、腸骨の下と陰毛の後ろに上下に伸びていました。一部の恐竜では、トカゲのように陰毛が前後に伸びていました。これが、シーリーがそれらを竜盤類、または「トカゲ寄棟」恐竜と呼んだ理由です。他の恐竜では、鳥の場合と同様に、陰毛が上下に伸び、イスキアの下を平行に走っていました。シーリーはこれらの恐竜を鳥盤類、または「鳥寄棟」恐竜と呼んだ。
原始的な竜盤類の骨盤の進化的変化から発達した鳥盤類の恐竜の骨盤(複数形の骨盤)。鳥盤類には、それらをグループ化する他の特徴もありました。前歯骨と呼ばれる骨が下顎の前にありました。また、「まぶた」の骨が眼窩の上部を縁取っていました。

一部の恐竜は竜盤類でも鳥盤類でもありませんでした。スタウリコサウルスやヘレラサウルスなど、最も初期の最も原始的な恐竜は、どちらの順序にも当てはまりません。彼らはあまりにも専門的で、恐竜の直接の祖先にはなれませんでした。
- 竜盤類:竜脚形亜目
- 竜盤類:獣脚類
- 鳥盤類:Thyneophora
- 鳥盤類:Marginocephalis
- 鳥盤類:鳥脚類
竜盤類:竜脚形亜目

竜脚形亜目はすぐに2つの主要なグループ、ProsauropodaとSauropodaに進化しました。竜脚類は以前に出現しましたが、既知の竜脚類は竜脚類の祖先ではなかった可能性があります。プロサウロポッドは広く普及しており、少なくとも7つの家族がいました。彼らは前期ジュラ紀まで生きました。最大の原竜脚類は、40フィート以上の長さのものもあり、いくつかの点で後の竜脚類に似たまっすぐな脚の恐竜でした。すべてのプロサウロポッドは植物を食べる人でした。
家族:Thecodontosauridae:最も原始的な原竜脚類であるThecodontosaurusも、最も小さいものの1つでした。長さは約6〜10フィートでした。すべての竜脚類およびほとんどの竜脚類と同様に、前足には目立つ爪があり、後足には大きな爪がありました。
家族:原竜脚類:これはプロサウロポッドの最も有名な家族であり、ヨーロッパ、中国、南北アメリカで動物が見られます。それらは25から30フィートの長さで、狭くて長い鼻、長い首、強力な前肢と後肢、そして重い体を持っていました。
I nfraorder:竜脚類
竜脚形亜目の2番目のグループである竜脚類は、おそらくテコドントサウルスによく似た祖先から来ました。これはおそらく、竜脚類が最初に出現した三畳紀後期に起こったと思われます。すべての竜脚類は巨人であり、4本足の植物を食べる人でした。
今日の象のように、竜脚類はその大きさのために捕食者をほとんど恐れていませんでした。大きいことはまた、彼らが木のてっぺんの葉など、小さな植物を食べる人には高すぎる食べ物に到達するのを助けました。竜脚類はサイズが大きいため、多くの特徴がありました。彼らは前足の把持機能を失い、足は長く真っ直ぐな柱のように見えました。彼らの椎骨(脊椎の骨)には、背骨の重量を軽くするための深いくぼみがありました。また、強度を高めるために、骨盤と脊椎が結合する椎骨が増えました。
竜脚類の頭蓋骨は鈍い(平らな)か先細になっている(ある点に来た)かのどちらかであり、鼻孔は鼻の先端から戻っていました。軽く造られて壊れやすい竜脚類の頭は、しばしば死後に壊れました。
家族:Vulcanodontidae:最も初期の真の竜脚類はジンバブエの前期ジュラ紀からのVulcanodonです。唯一の骨格には、頭、首、そして尾の多くが欠けています。体はかさばり、足は長く真っ直ぐでした。前肢は後肢とほぼ同じ長さで、各後足には5本のつま先がありました。
家族:バラパサウルス科:次に最も原始的な竜脚類であるバラパサウルスは、インドの前期ジュラ紀のいくつかの骨格の一部から知られています。長さが最大60フィートで、体が細く、首、尾、手足が長い場合。

家族:Euhelopodidae:中国の中期および後期ジュラ紀から知られている竜脚類のほとんどは、現在、別の家族、Euhelopodidaeに配置されています。Euhelopodidsは、より原始的な竜脚類の家族の1つですが、非常に首の長いマメンチサウルスやオメイサウルスなどのエキゾチックな動物が含まれています。
家族:ケティオサウルス科:この家族は中期ジュラ紀から、おそらくヴルカノドン科の祖先からです。ケティオサウルスは、中期ジュラ紀によってヨーロッパ、南北アメリカ、アフリカ、オーストラリアに拡大し、広がりました。
ケティオサウルスの頭蓋骨は鈍く箱のようで、鼻の側面に鼻孔がありました。首は短く、通常は12個の椎骨がありました。竜脚類の場合、それらは小さいものから大きいものまでありました。ほとんどは35から60フィートの長さでした。最も有名な属は、中国の中期ジュラ紀のシュノサウルスです。尻尾の先に小さな骨のあるクラブがありました。ケティオサウルスはジュラ紀後期まで続きました。
家族:ブラキオサウルス科:ブラキオサウルスの前肢は後肢と同じかそれより長かった。これにより、体は首から尻尾に向かって後方に傾斜しました。この家族の首の椎骨の数は13以上に増加しました。鼻孔は、鈍い鼻の先端からさらに後ろにあり、ブラキオサウルスの頭蓋骨の目の上にありました。ほとんどのブラキオサウルスはケティオサウルスよりも大きく、尾は短くても80フィート以上の長さでした。彼らは知られている中で最も重い陸上動物の1つでした。体重を減らすために、彼らの巨大な脊椎はほぼ完全に中空でした。世界的に知られているブラキオサウルスは、中期ジュラ紀の化石記録に現れ、ジュラ紀後期に最も多く、前期白亜紀の終わりまでにほとんど消滅しました。
家族:カマラサウルス科:この家族では、頭蓋骨は箱のようでした。彼らはまだ12の首の椎骨を持っていて、前肢は後肢よりわずかに短かった。最後に知られているカマラサウルスの1つは、モンゴルのオピストコエリカウディアでした。これは、尾が短い重い体の竜脚類で、後肢に立って餌を食べるときにそれを支えるのにおそらく役立ちました。
家族:ディプロドクス科:この家族には、アパトサウルスやディプロドクスなど、最もよく知られている竜脚類がいくつか含まれています。ディプロドクスの頭蓋骨は長く、スプーンの形をした鼻に向かって先細になっており、頭蓋骨の上に鼻孔がありました。彼らの小さな棒状の歯は彼らの鼻の前にありました。ディプロドクスは首が長く、最大15個の椎骨がありました。彼らの背中は後肢の長さに比べて短く、尻尾はおそらく武器として使用されたむち打ち症で終わっていました。長い首と尾は、いくつかのディプロドクスをこれまでに生きた中で最も長い動物にしました。
家族:ティタノサウルス科:白亜紀後期からのほとんどすべての南半球の竜脚類、および多くの初期のものは、チタノサウルスでした。彼らの手足はずんぐりしていた。尾の前部と中央からの椎骨は独特であり、家族を区別する最高の特徴です。完全またはほぼ完全なチタノサウルスの頭蓋骨は1つも見つかりませんでした。最も興味深いティタノサウルスの1つはサルタサウルスでした。サルタサウルスはしゃがんでいて、曲竜類に似た鎧で覆われていました。
ほとんどのティタノサウルスは約40〜50フィートの長さでしたが、いくつかは巨大になりました。ティタノサウルスは白亜紀に主に南半球に生息し、北半球の竜脚類が絶滅したため、そこで生き残った。
竜盤類:獣脚類

獣脚類は、herrerasauriansを除くすべての略奪的な恐竜でした。最小の恐竜から最大の肉食動物まで、獣脚類はすべてのサブオーダーの中で最も異なる種類の竜盤類の恐竜を持っていました。これらの二本足の肉食者は、3つ以下の機能的なつま先で足を爪でつめていました。
鳥の羽と足は獣脚類の骨格の腕と足に似ています。また、鳥のように、すべての獣脚類はある程度中空の骨を持っていました。最高の祖先の鳥は、ジュラ紀後期の小さな羽毛のある始祖鳥のような始祖鳥です。
獣脚類は2つの主要なグループに進化しました。柔軟な尾を持つケラトサウルス。尾が硬いテタヌラ類。初期の獣脚類はすべてceratosauriansでした。彼らの化石記録は、三畳紀後期からジュラ紀後期までのものです。テタヌラ類は中期ジュラ紀に出現し、後期ジュラ紀に多様化し、白亜紀後期まで北半球の主要な捕食者でした。
インフラオーダー:ケラトサウルス
インフラオーダー:ケラトサウルス
家族:ポドケサウリダエ:初期のセラトサウルスには、北アメリカ西部の後期三畳紀のコエロフィシスが含まれます。それは小さくて機敏で、多くの歯を持つ長くて細い頭蓋骨を持っていました。
家族:ハルティコサウルス科とケラトサウルス科:前期ジュラ紀に生息していたディロフォサウルスは、頭に二重の紋章がありました。ケラトサウルスはジュラ紀後期のもので、頭に角がありました。どちらも北米出身で、後のセラトサウルスのメンバーの例です。ジュラ紀後期以降、セラトサウルス人は北半球で姿を消したようですが、南アメリカでは生き残りました。
家族:アベリサウルス科:アベリサウルスは、短くて背の高い頭蓋骨を特徴とする中型から大型のアフリカおよび南アメリカの獣脚類のグループです。アルゼンチンのカルノタウルスとマダガスカルのマジュンガサウルスは、カルノタウルスの頭蓋骨に2つの大きな角があることを除けば、似ています。

I nfraorder:Tetanurae
最先端の獣脚類であるテタヌラ類には、関係がよく理解されていないいくつかのグループが含まれていました。頭の上の紋章や他の装飾は通常存在しませんでした。彼らの手は3本以下の指を持っていて、「親指」は通常最大の爪を持っていました。
家族:コンプソグナトゥス科:最も原始的なテタヌラ類は、ヨーロッパの後期ジュラ紀のコンプソグナトゥスでした。それは最小の獣脚類で、長さ約3フィート、軽く造られていました。
家族:Coeluridae:北米西部でのジュラ紀後期の間に住んでいたオルニトレステスとコエルルスは、腰に2〜3フィートであり、長い10フィートまでの6から軽く建て獣脚類、高速実行していました。
家族:カルカロドントサウルス科:ゴンドワナ大陸の巨大な獣脚類のこのグループには、巨大な略奪恐竜、アルゼンチンのギガノトサウルス、北アフリカのカルカロドントサウルスが含まれます。
家族:テリジノサウルス科:テリジノサウルスは、アジアと北アメリカの後期白亜紀から知られている明らかに草食性または雑食性の獣脚類でした。テリジノサウルスの珍しい鳥のような骨盤とほぼプロサウロポッドのような頭蓋骨は、それらの進化の位置について不確実性をもたらしましたが、最近、それらはオルニトミムシ科に密接に関連する獣脚類であることが示されました。
家族:スピノサウルス科:スピノサウルスは、背中に帆のような構造を形成した可能性のある、長いワニのような鼻と細長い脊椎の棘を持つ獣脚類の特徴的なグループです。スピノサウルスは白亜紀に限定されていますが、アフリカ、南アメリカ、ヨーロッパで知られています。
家族:Oviraptorisauridae:アジアの白亜紀からのもう一つの好奇心が強い獣脚類はOviraptorであり、それはカメのようなくちばしを持つ背の高い、非常に空気圧の頭蓋骨を持っています。オヴィラプトルは、もともとケラトプスの卵の巣であると考えられていた標本がモンゴルで発見されたため、その名前(「卵捕食者」)が付けられました。
家族:アロサウルス科:この家族は、15〜35フィート以上の長さのより大きなジュラ紀および前期白亜紀の獣脚類の典型です。最大のアロサウルスは40フィート以上の長さだったかもしれません。アロサウルスは細身ですが危険な捕食者でした。
家族:ティラノサウルス科:すべての獣脚類の中で最も劇的なのは、おそらく白亜紀後期の初めにアロサウルスのような祖先から来たティラノサウルスでした。他のテタヌラ類とは異なり、彼らは巨大な体、異常な形の頭、そして小さな2本の指の手を持っていました。

いくつかのティラノサウルスは50フィート近くの長さで、知られている中で最大の肉食性の陸上動物になりました。ナノティラヌスなどの最小のものは、約18フィートの長さでした。アルバートサウルスやダスプレトサウルスなどの中型のティラノサウルスは、長さが約30フィートでした。
家族:オルニトミムシ科:白亜紀にも多くの種類の小さな獣脚類が発生しました。彼らの骨格は鳥のようでした。ほとんどの人は、強力な打撃アクションのために前脚と手に変更がありました。前肢の「クイックストライク」モーションは、鳥の羽のパワーストロークの始まりだった可能性があります。
「ダチョウの恐竜」またはOrnithomimidaeは、東アジアおよび北アメリカ西部の白亜紀後期から最もよく知られています。彼らは頭が小さく、通常は歯がありませんでした。彼らは長い首と短くて硬い背中を持っていました。彼らの前肢は長く、彼らの強力な後脚はスピードのために作られました。彼らは最速の恐竜だったと考えられています。
家族:ドロマエオサウルス科:デイノニクスやその他の「鎌の爪」の獣脚類は、最もよく研究されている恐竜の1つです。デイノニクスの発見は、鳥と恐竜の関係のアイデアを支持し、恐竜が温血動物であるという議論を開始しました。各足には、第2趾に大きな鎌形の爪がありました。尾の端には椎骨があり、椎骨が互いに固定されて硬くなっていました。
家族:トロオドン科:この「鎌の爪」の獣脚類のグループは、大きな脳と大きな目が前を向いていました。トロオドンやサウロルニトイデスなどの一部は、一部の哺乳類とほぼ同じくらい頭が良いかもしれません。
鳥盤類:Thyneophora

初期の鳥盤類恐竜は、アルゼンチンの中期三畳紀後期の長さ3フィート、2本足(二足歩行)の植物を食べるピサノサウルスでした。すべての鳥盤類は植物を食べる人でした。
その後、鳥盤類は3つの高度なグループに分かれました。四つん這いで歩く重くて装甲のある植物を食べる人。特殊なドーム型の恐竜と角のある恐竜。そして、イグアノドンとアヒルの子恐竜を含む二本足の植物を食べる人。
家族:ファブロサウルス科:この家族は、いくつかの大陸の三畳紀後期からジュラ紀後期の岩石に見られます。他の原始的な鳥盤類は通常この家族に分類されます。南アフリカの前期ジュラ紀のファブロサウルスにちなんで名付けられました。すべてのファブロサウルスは小さな二足歩行の植物を食べる人でした。最も有名なファブロサウルスは、同じくアフリカ南部の前期ジュラ紀のレソトサウルスです。レソトサウルスの骨盤は、鳥盤類の底にそれを置くいくつかの特徴を示しています。北アメリカ西部の前期ジュラ紀のスクテロサウルスは、後の装甲鳥盤類の大きなプレートに似た小さな骨プレートによって保護されていました。
亜目:装盾亜目
インフラオーダー:ステゴサウリア
剣竜類はジュラ紀の主な装甲恐竜でした。曲竜類は背景に残った。しかし、白亜紀には曲竜類が剣竜類に取って代わった。スケリドサウルスは両方のグループに非常によく似ていました。それはイギリスの前期ジュラ紀からのものであり、四肢すべて(四肢)を歩いていました。それは、ステゴサウルスのような歯、装甲板と棘のアンキロサウルスのようなパターン、そしてスクテロサウルスのような骨盤を持っていました。
家族:ステゴサウルス科:これらの装甲恐竜は、おそらく前期ジュラ紀の間に中国で進化しました。ジュラ紀後期までに、彼らはヨーロッパ(ダケントルルス)、北アメリカ(ステゴサウルス)、アフリカ(ケントロサウルス)にいました。
鎧は、頭の後ろから尻尾まで背中に沿って走る2列の大きな骨のプレートでした。尾の端にある鋭い棘は、捕食者に対する武器として使用されました。
下目:曲竜類
白亜紀初期には、インドを除くすべての場所で曲竜類が剣竜類に取って代わりました。曲竜類は、2列の背の高いプレートではなく柔軟なボディアーマーを備えていたため、剣竜類とは異なりました。彼らはまた地面に近く、背中にわずかなアーチがあったとしてもわずかでした。剣竜類の頭は長くて狭いが、曲竜類は短い板状の頭蓋骨を持っていた。尾棘の代わりに、曲竜類は防御のために肩棘または尾棘を持っていました。
家族:ノドサウルス科:すべてのジュラ紀および南半球の属を含む、より原始的な曲竜類がこの家族に属しています。いくつかのノドサウルスは、保護のために首と肩に沿って大きな円錐形の棘を持っていました。
家族:アンキロサウルス科:この家族は、白亜紀初期にノドサウルスの祖先から生まれた可能性があります。アンキロサウルスの頭蓋骨は後ろから角が突き出ており、上から見ると三角形になっています。すべてのアンキロサウルスには、防御のための巨大な骨の尾のクラブがありました。
鳥盤類:Marginocephalis

イングランドの前期白亜紀のヤヴェルランディアは、最も古くから知られている堅頭竜類です。しかし、唯一知られている標本は、動物がどのように見えたか、またはその関係についてほとんど示さない、2つの骨の塊を持つ小さくて厚い頭蓋骨です。ドーム型の恐竜は白亜紀の希少な中型から中型の動物でした。ほとんどが北半球に住んでいましたが、マジュンガサウルスという1つの属は、マダガスカルの後期白亜紀から知られています。
ドーム型の恐竜の後頭部は、しばしば骨の塊や短いスパイクのある棚に広げられました。1つの家族、ホマロケファレ科は、頭蓋骨の上部に平らで厚い骨を持っていました(モンゴルのホマロケファレとゴヨケファレのように)。
他の家族である堅頭竜類では、骨は非常に厚くて高いドームに隆起しており、これが動物の外観の主な特徴であり、棚の上にも成長していました(ステゴケラスのように)。パキケファロサウルスの骨格はまれですが、頭蓋骨のドームは固い骨であるため、化石化することがよくありました。それらはいくつかの場所で非常に一般的です。
堅頭竜類は広くてぽっちゃりした体を持っていました。彼らは二足歩行の植物を食べる人でした。動物が成長するにつれて、そのドームは大きくなりました。厚いドームは、オス同士の頭突きコンテストで仲間を競ったり、捕食者を横に突き合わせて戦ったりするために使用されました。
下目:角竜類
角竜類は角竜類とその親戚でした。それらは、大きなオウムのようなくちばしの一部を形成する特別な骨、吻側を持っていたため、堅頭竜類(および他のすべての恐竜)とは異なりました。
家族:プシッタコサウルス科:最も古く、最も原始的な角竜類は、中国とモンゴルの前期白亜紀からのプシッタコサウルスなどの小さな二足歩行のランナーのこの家族に属しています。
家族:プロトケラトプス科:この家族では、頭蓋骨の後ろが首の後ろの広いフリルに拡張されました。ミクロケラトゥスは、その可能性のあるpsittacosauridの祖先のように、二足歩行の動物でした。他のプロトケラトプスは四つん這いで歩き、大きな頭を支えやすくしました。
家族:ケラトプス科:ケラトプス科は、恐竜の家族の中で最も短い範囲を持っていました。それらは北アメリカ西部の白亜紀後期に発生しました。彼らはすぐに多くの珍しい形に進化し、中生代の終わりまで生きました。牛から象のサイズまで、四足(4本足)のケラトプスは頭に角とフリルがありました。

彼らは丈夫な植物をスライスするための何百もの歯を持つ強力な顎を持っていました。トリケラトプスは、陸生動物の中で最も強力な顎の筋肉を持っていました。角は武器として使われました。フリルは首を保護していて、交尾期に向けて鮮やかな色になっている可能性があります。フリルはまた、体温を均一に保つのに役立った可能性があります。
鳥盤類:鳥脚類

サイズの増加とすべての恐竜の最も顕著な咀嚼配置の進化を除いて、異なる鳥脚類の家族は細部が互いに異なっていました。全員が二足歩行の植物を食べる人でした。
家族:ヘテロドントサウルス科:これらの小さくて軽快な二足歩行の植物を食べる人は、主にアフリカ南部の前期ジュラ紀の岩石で発見されています。
彼らの歯は前が鋭くて牙のようでしたが、顎の側面の歯は植物を噛んだりスライスしたりするために作られました。上顎と下顎の角に大きな犬のような歯(円錐形の尖った歯)がありました。
家族:ヒプシロフォドン科:これは最も広範で最も寿命の長い鳥脚類の家族でした。それは中期ジュラ紀から白亜紀の終わりまでほぼ世界中で繁栄しました。
ヒプシロフォドンチドは小さかったが、頭は比較的大きかった。彼らの足は4つの機能的なつま先で原始的でした。最初のつま先は他のつま先よりも小さかった。第5趾は副子だけでした。ヒプシロフォドンチドは、小さな手で小さな前肢を持っていました。骨の腱は背中を強化し、尻尾を硬くしました。ほとんどは長さ約6〜10フィートの小さな恐竜でしたが、一部のテノントサウルス種は22フィートもの長さでした。白亜紀後期のテスケロサウルスは約18フィートの長さに成長しました。
家族:ドリオサウルス科:この短命の家族は、ヒプシロフォドン科とほぼ同じ時期に発生しました。最も初期のドリオサウルスは、北アメリカ西部とアフリカ東部のジュラ紀後期のドリオサウルスでした。ヨーロッパと北アフリカの前期白亜紀のヴァルドサウルスとアフリカのカングナサウルスは、この家族の他の2つの属です。前肢と頭が小さいドライサウリドは、ヒプシロフォドンチドよりも大きく、より強力でした。彼らは鼻の前に歯がなく、代わりに角質の覆いを持っていたかもしれないよく発達したくちばしを持っていました。
家族:カンプトサウルス科:北アメリカ西部のジュラ紀後期のカンプトサウルス属は、長さ約15フィートのぽっちゃりした中型の鳥脚類でした。それは特殊な足と頭蓋骨を持っていました。
家族:イグアノドン科:イグアノドンは最も有名な恐竜の1つです。このかさばる、長さ35フィートの鳥脚類は、深くて狭い頭蓋骨を持っていました。強く、よく発達した骨盤; 背中に沿って走る骨の腱の列。親指が鋭いスパイクになった手。3本の広いつま先と内側のつま先がスプリントになりました。歯は厚く、常に交換されていました。丈夫な植物を食べ、イグアノドンは両足で歩きましたが、手を使って四つんばいで歩くこともできました。

Iguanodontidsは、北アメリカ、北アフリカ、ヨーロッパ、およびアジアの白亜紀初期および後期の地層で発見されています。最も進んだのは中央アジアのプロバクトロサウルスと北アフリカのオウラノサウルスでした。プロバクトロサウルスの歯は、イグアノドンの歯よりも複雑な置換パターンを持っていました。オウラノサウルスの椎骨は非常に長い棘を持っていて、帆を作りました。アヒルの子恐竜の台頭に伴い、イグアノドンチドは衰退しました。
家族:ハドロサウルス科:このグループは、ハドロサウルス科とランベオサウルス科の2つのグループで構成されています。それらは両方とも密接に関連したアヒルの子恐竜でした。彼らは、南北アメリカとユーラシアの白亜紀後期の大規模から非常に大規模な植物を食べる人でした。最大のものは中国のシャントゥンゴサウルスとメキシコのランベオサウルス・ラティコーダスで、どちらも長さ50フィート、体重20トンを超えた可能性があります。彼らは両足で歩くことができる最大の既知の動物だったかもしれません。ほとんどのアヒルの子恐竜は約30フィートの長さでした。
歯は動くエスカレーターの階段のように30から40の垂直の列にありました。それぞれの歯がすり減ったので、それはその真下の歯に置き換えられました。このプロセスは、動物が孵化したとき(またはその前でさえ)に始まり、生きている限り続きました。ハドロサウルスの頭蓋骨は一般的に長く、ランベオサウルスの頭蓋骨ほど深くはなく、アヒルのようなくちばしはより平らで幅が広かった。
いくつかのハドロサウルスは頭蓋の頂上を欠いていました(エドモントサウルス)。他の人は鼻骨をアーチ状にした(クリトサウルス)。他の人はしっかりした頭蓋の頂上を持っていました(プロサウロロフスとマイアサウラ)。しかし、頭蓋骨がよく知られているすべてのランベオサウルス(ヒパクロサウルス、コリトサウルス、ランベオサウルス、パラサウロロフス)には、音を出すために使用された可能性のあるループ状の鼻腔を備えた中空の頂上がありました。
アヒルの子恐竜はおそらくすべての恐竜の中で最も進んだものでした。彼らは優れた聴覚、視力、声、そして嗅覚を持っていました。彼らは巨大な群れに住んでいて、季節ごとに移動し、毎年同じ場所に戻って交尾し、産卵した可能性があります。